
Gentian Violet Inhibits Cell Proliferation through Induction of Apoptosis in Ovarian Cancer Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Proliferation Assay
2.3. TUNEL Assay
2.4. Western Blot Analysis
2.5. Measurement of ROS Generation
2.6. Statistical Analyses
3. Results
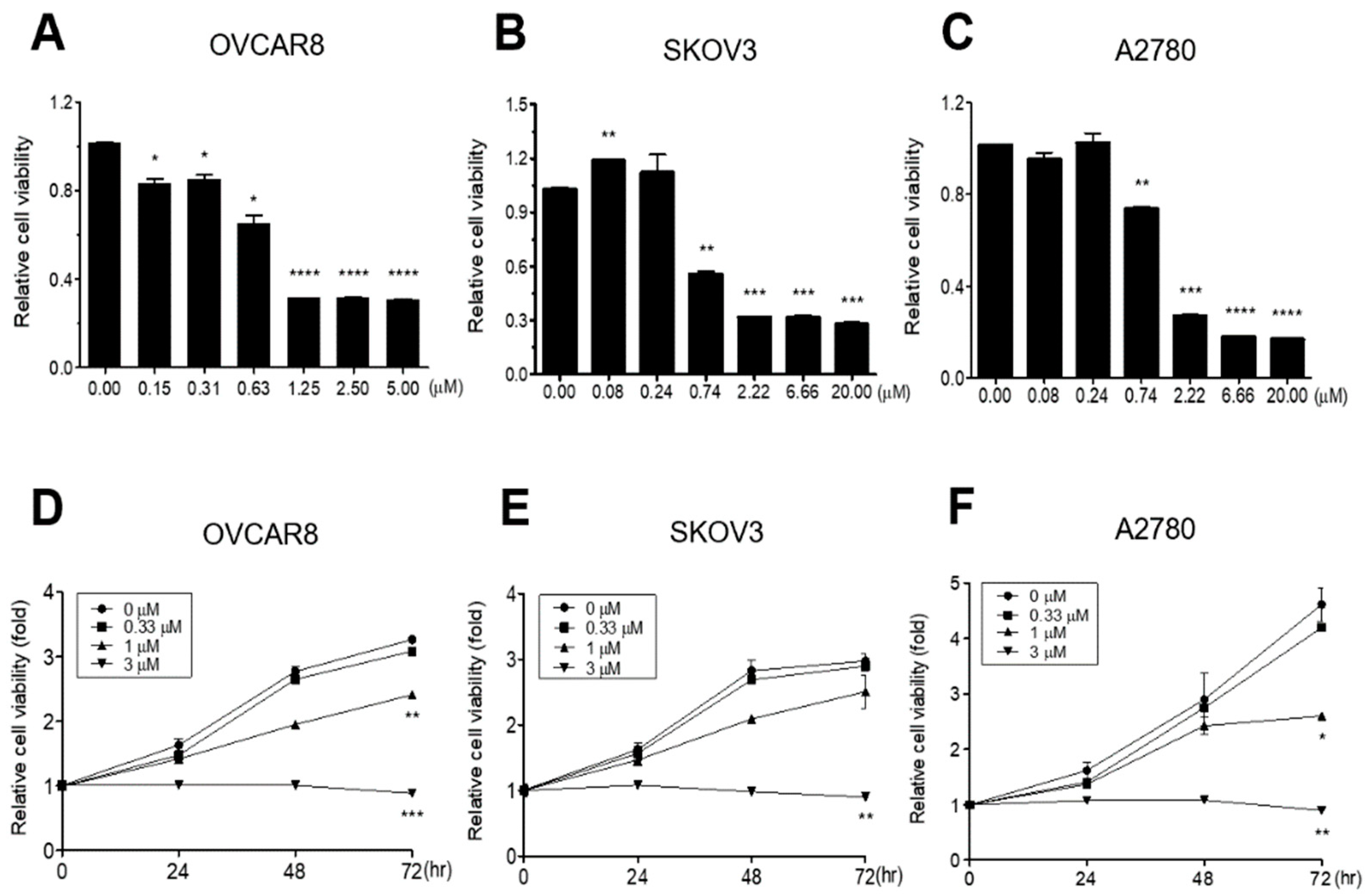
3.1. GV Inhibits Proliferation of OC Cells
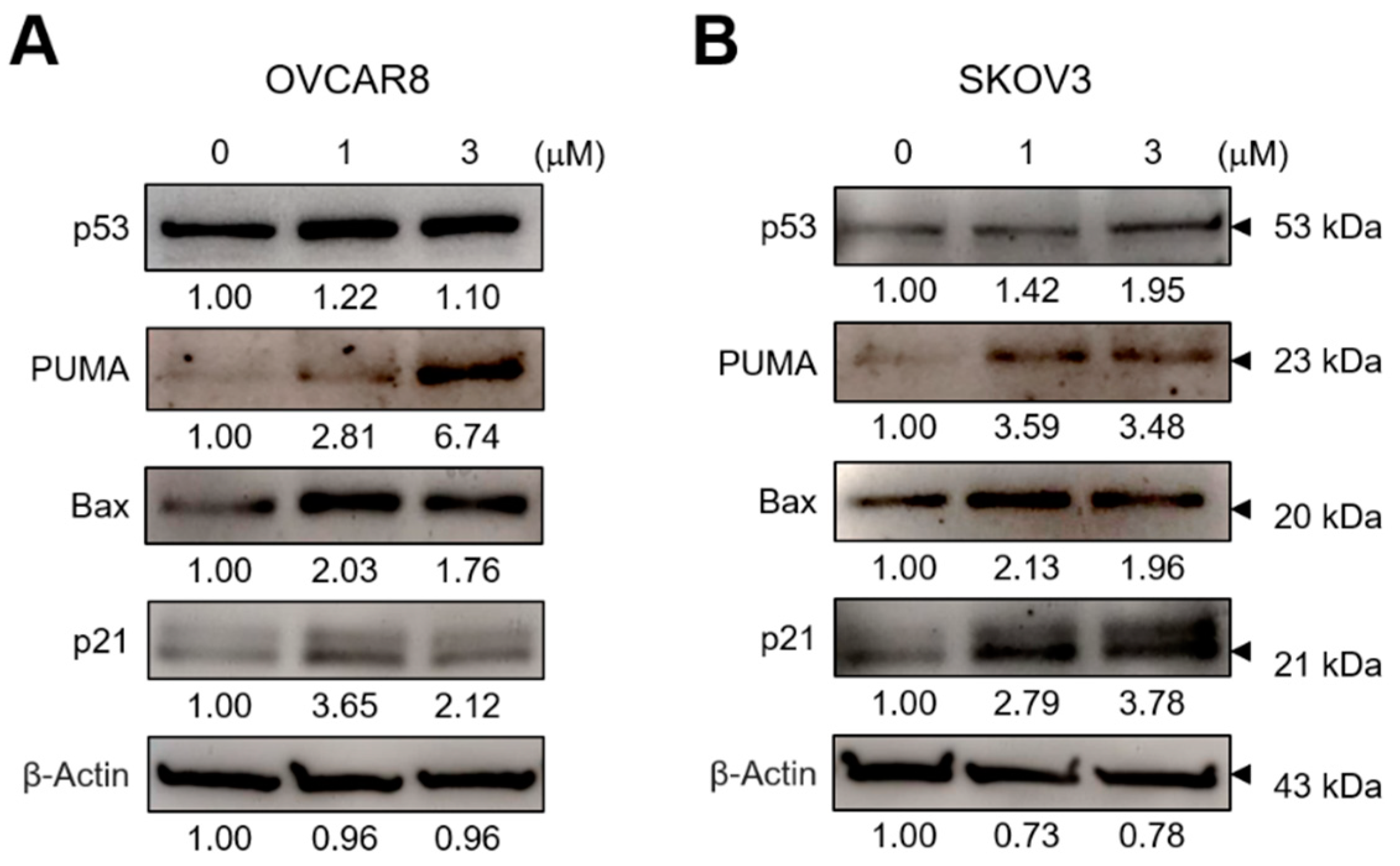
3.2. GV Induces Apoptosis in OC Cells
3.3. GV Increases ROS Levels in OC Cells
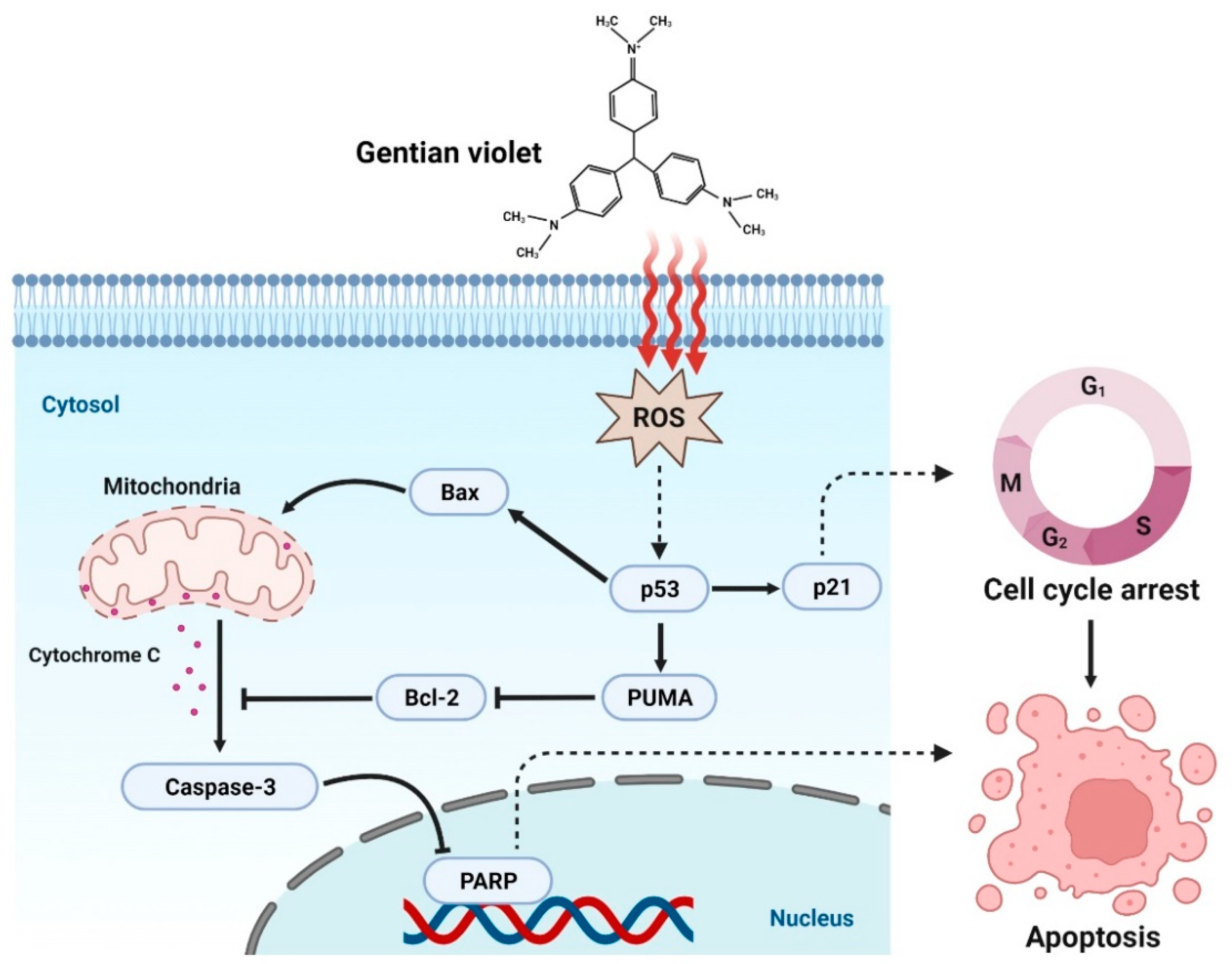
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in ovarian cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, B.T.; Coleman, R.L.; Markman, M. Ovarian cancer. Lancet 2009, 374, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurnit, K.C.; Fleming, G.F.; Lengyel, E. Updates and new options in advanced epithelial ovarian cancer treatment. Obstet. Gynecol. 2021, 137, 108–121. [Google Scholar] [CrossRef]
- Mao, C.L.; Seow, K.M.; Chen, K.H. The utilization of bevacizumab in patients with advanced ovarian cancer: A Systematic Review of the Mechanisms and Effects. Int. J. Mol. Sci. 2022, 23, 6911. [Google Scholar] [CrossRef]
- Sambasivan, S. Epithelial ovarian cancer: Review article. Cancer Treat. Res. Commun. 2022, 33, 100629. [Google Scholar] [CrossRef]
- Sato, S.; Itamochi, H. Bevacizumab and ovarian cancer. Curr. Opin. Obstet. Gynecol. 2012, 24, 8–13. [Google Scholar] [CrossRef]
- Mittica, G.; Ghisoni, E.; Giannone, G.; Genta, S.; Aglietta, M.; Sapino, A.; Valabrega, G. PARP inhibitors in ovarian cancer. Recent Pat. Anticancer Drug Discov. 2018, 13, 392–410. [Google Scholar] [CrossRef]
- Montoni, A.; Robu, M.; Pouliot, E.; Shah, G.M. Resistance to PARP-inhibitors in cancer therapy. Front. Pharmacol. 2013, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valabrega, G.; Scotto, G.; Tuninetti, V.; Pani, A.; Scaglione, F. Differences in PARP inhibitors for the treatment of ovarian cancer: Mechanisms of action, Pharmacology, Safety, and Efficacy. Int. J. Mol. Sci. 2021, 22, 4203. [Google Scholar] [CrossRef] [PubMed]
- Maley, A.M.; Arbiser, J.L. Gentian violet: A 19th century drug re-emerges in the 21st century. Exp. Dermatol. 2013, 22, 775–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabha, N.; Arora, R.D.; Ganguly, S.; Chhabra, N. Gentian violet: Revisited. Indian J. Dermatol. Venereol. Leprol. 2020, 86, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, F.; Yang, H.; Wen, J.; Tang, Y.; Wan, F.; Zhang, X.; Wu, J. Gentian violet induces apoptosis and ferroptosis via modulating p53 and MDM2 in hepatocellular carcinoma. Am. J. Cancer Res. 2022, 12, 3357–3372. [Google Scholar]
- Garufi, A.; D’Orazi, V.; Arbiser, J.L.; D’Orazi, G. Gentian violet induces wtp53 transactivation in cancer cells. Int. J. Oncol. 2014, 44, 1084–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrobono, S.; Morandi, A.; Gagliardi, S.; Gerlini, G.; Borgognoni, L.; Chiarugi, P.; Arbiser, J.L.; Stecca, B. Down-regulation of SOX2 underlies the inhibitory effects of the triphenylmethane gentian violet on melanoma cell self-renewal and survival. J. Investig. Dermatol. 2016, 136, 2059–2069. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wood, G.S. Analysis of the effect of gentian violet on apoptosis and proliferation in cutaneous T-cell lymphoma in an in vitro study. JAMA Dermatol. 2018, 154, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Vikulina, T.; Weitzmann, M.N. Gentian violet inhibits MDA-MB-231 human breast cancer cell proliferation, and reverses the stimulation of osteoclastogenesis and suppression of osteoblast activity induced by cancer cells. Oncol. Rep. 2015, 34, 2156–2162. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Fierro, A.; Duenas-Gonzalez, A. Drug repurposing for cancer therapy, easier said than done. Semin. Cancer Biol. 2021, 68, 123–131. [Google Scholar] [CrossRef]
- Wurth, R.; Thellung, S.; Bajetto, A.; Mazzanti, M.; Florio, T.; Barbieri, F. Drug-repositioning opportunities for cancer therapy: Novel molecular targets for known compounds. Drug Discov. Today 2016, 21, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Beaufort, C.M.; Helmijr, J.C.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van IJcken, W.F.; Heine, A.A.; Smid, M.; et al. Ovarian cancer cell line panel (OCCP): Clinical importance of in vitro morphological subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.K.; Davis, D.A.; Tomar, S.; Roy, L.; Gurler, H.; Xie, J.; Lantvit, D.D.; Cardenas, H.; Fang, F.; Liu, Y.; et al. In vivo tumor growth of high-grade serous ovarian cancer cell lines. Gynecol. Oncol. 2015, 138, 372–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Perry, B.N.; Govindarajan, B.; Bhandarkar, S.S.; Knaus, U.G.; Valo, M.; Sturk, C.; Carrillo, C.O.; Sohn, A.; Cerimele, F.; Dumont, D.; et al. Pharmacologic blockade of angiopoietin-2 is efficacious against model hemangiomas in mice. J. Investig. Dermatol. 2006, 126, 2316–2322. [Google Scholar] [CrossRef]
- Brandes, R.P.; Weissmann, N.; Schroder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef]
- Kleniewska, P.; Piechota, A.; Skibska, B.; Goraca, A. The NADPH oxidase family and its inhibitors. Arch. Immunol. Ther. Exp. 2012, 60, 277–294. [Google Scholar] [CrossRef]
- Xia, C.; Meng, Q.; Liu, L.Z.; Rojanasakul, Y.; Wang, X.R.; Jiang, B.H. Reactive oxygen species regulate angiogenesis and tumor growth through vascular endothelial growth factor. Cancer Res 2007, 67, 10823–10830. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, T.; Nakagawara, A. Role of p53 in Cell Death and Human Cancers. Cancers 2011, 3, 994–1013. [Google Scholar] [CrossRef] [Green Version]
- Stein, Y.; Rotter, V.; Aloni-Grinstein, R. Gain-of-function mutant p53: All the roads lead to tumorigenesis. Int. J. Mol. Sci. 2019, 20, 6197. [Google Scholar] [CrossRef] [Green Version]
- Puca, R.; Nardinocchi, L.; Starace, G.; Rechavi, G.; Sacchi, A.; Givol, D.; D’Orazi, G. Nox1 is involved in p53 deacetylation and suppression of its transcriptional activity and apoptosis. Free Radic. Biol. Med. 2010, 48, 1338–1346. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.S.; Kim, J.H.; Lee, C.Y.; Lee, Y.M.; Lee, S.; Chang, H.K.; Kim, H.J.; Heo, K. Gentian Violet Inhibits Cell Proliferation through Induction of Apoptosis in Ovarian Cancer Cells. Biomedicines 2023, 11, 1657. https://doi.org/10.3390/biomedicines11061657
Choi MS, Kim JH, Lee CY, Lee YM, Lee S, Chang HK, Kim HJ, Heo K. Gentian Violet Inhibits Cell Proliferation through Induction of Apoptosis in Ovarian Cancer Cells. Biomedicines. 2023; 11(6):1657. https://doi.org/10.3390/biomedicines11061657
Chicago/Turabian StyleChoi, Min Sung, Ji Hyeon Kim, Chae Yeon Lee, Yul Min Lee, Sukmook Lee, Ha Kyun Chang, Hyun Jung Kim, and Kyun Heo. 2023. "Gentian Violet Inhibits Cell Proliferation through Induction of Apoptosis in Ovarian Cancer Cells" Biomedicines 11, no. 6: 1657. https://doi.org/10.3390/biomedicines11061657